
Цягліцы

Цягліцы[1][2] (па-лацінску: musculi) — органы цела жывёлаў і чалавека, якія складаюцца з пругкай элястычнай цяглічнай тканкі, якая здольная скарачацца пад уплывам нэрвовых імпульсаў. Яны прызначаныя да розных мэтаў, як то перамяшчэньне цела ў прасторы, рух галавы, хрыбетніка й вачэй, праца канцавінаў і міміка(en)[3], скарачэньне галасавога вязу й дыханьня. Цягліцы на 86,3 адсоткаў складаюцца з вады.
Цягліцы дазваляюць рухаць часткамі цела й адлюстраваць у дзеяньні думкі й пачуцьці. Чалавек выконвае любыя рухі — ад такіх простых, як плюсканьне й усьмешка, да тонкіх і жвавых, якія можна заўважыць у дзеяньнях ювэліраў і спартоўцаў — дзякуючы здольнасьцям цягліцавых тканак скарачацца. Ад спраўнай працы цягліцаў, якія складаюцца з трох групаў, залежыць ня толькі рухомасьць арганізму, але й функцыянаваньне ўсіх фізыялягічных працэсаў. Працай усіх цягліцавых тканак кіруе нэрвовая сыстэма, якая забясьпечвае іхную сувязь з галаўным мозгам і млечам і рэгулюе пераўтварэньне хімічнай энэргіі ў мэханічную.
У целе чалавека налічваецца вялікая колькасьць цягліцаў, толькі шкілетавых зь іх маецца ў целе чалавека каля 600[4]. Самыя маленькія прымацаваныя да маленькіх косткаў, якія зьмяшчаюцца ў вуху. Самыя вялікія — крумянковыя цягліцы, якія адказваюць за рухомасьць нагі. Самыя дужыя цягліцы — жавальныя. Паводле формы цягліцы вельмі разнастайныя. Часьцей за ўсё сустракаюцца пачынкавападобныя цягліцы, характэрныя для канцавінаў, і шырокія цягліцы — яны ўтвораць сьценкі тулава. Калі ў цягліцаў агульнае сухажыльле, а галовак дзьве або больш, то іх называюць двух-, трох- або чатырохгаловымі цягліцамі. Цягліцы й шкілет вызначаюць форму чалавечага цела. Актыўны лад жыцьця, збалянсаванае харчаваньне й занятак спортам спрыяюць разьвіцьцю цягліцаў і памяншаюць аб’ём тлушчавай тканіны.
Анатомія
[рэдагаваць | рэдагаваць крыніцу]Анатомія цягліцаў улучае ў сябе як тапаграфічную анатомію, якая ўключае ў сябе ўсе цягліцы арганізму, а, з другога боку, гісталёгію, якая ўключае ў сябе структуры адной цягліцы.
Тыпы тканак
[рэдагаваць | рэдагаваць крыніцу]Цягліцавая тканка зьяўляецца мяккай тканкай, і ёсьць адным з чатырох асноўных тыпаў тканкаў у жывёлаў, аднак толькі тры тыпы цягліцавых тканкаў налічваецца ў хрыбетных, як то шкілетныя цягліцы, гладкія цягліцы й сардэчная цягліца ці міякард.
Шкілетныя цягліцы
[рэдагаваць | рэдагаваць крыніцу]

Шкілетныя цягліцы або «добраахвотныя цягліцы», якія мацуюцца да костак з дапамогай сухажыльляў альбо апанэўрозаў у некалькіх месцах да косткі й выкарыстоўваюцца для ажыцьцяўленьня шкілетнага руху, як то перамяшчэньне й падтрыманьне паставы. Нягледзячы на тое, што пастуральны кантроль, як правіла, падтрымліваецца як несьвядомы рэфлекс, цягліцы адказныя рэагаваць на сьвядомы кантроль як непастуральныя цягліцы. У сярэдняга дарослага мужчыны шкілетныя цягліцы складаюць 42% ад масы цела, у той час як у жанчыны гэты паказьнік роўны 36%[5]. З дапамогай спэцыяльных практыкаваньняў памер цягліцавых вузаў можна павялічваць да тых часоў, пакуль яны не вырастуць у масе й аб’ёме й ня стануць рэльефнымі. Скарачаючыся, цягліца кароціцца, патаўшчаецца й рухаецца адносна суседніх цягліцаў. Укарачэньне цягліцы суправаджаецца збліжэньнем ейных канчараў і костак, да якіх яна прымацоўваецца. У кожным руху ўдзельнічаюць як цягліцы, якія ажыцьцяўляюць рух, гэтак і процідзейныя яму цягліцы, што надае руху дакладнасьць і плыўнасьць.
Шкілетныя цягліцы падзяляюцца на дзьве шырокія катэгорыі, то бок якія павольна скарачаюцца й хутка скарачаюцца.
Цягліцы першага тыпу, якія павольна скарачаюцца, ці «чырвоныя цягліцы», шчыльна ўсеяныя капілярамі й багатыя на мітахондрыю й міяглябін, што надае цягліцавай тканцы характэрны чырвоны колер. Такія цягліцы могуць несьці больш кіслароду й падтрымліваюць аэробную актыўнасьць праз выкарыстаньне тлушчаў альбо вугляводаў у якасьці паліва[6]. Гэтыя цягліцы здольны скарачацца на працягу працяглых пэрыядаў часу, але зь невялікай сілай.
Хуткаскарачальныя цягліцы маюць тры асноўныя падтыпы, якія адрозьніваюцца скарачальнай хуткасьцю[7] й сілай[6]. Цягліцы гэтага тыпу зьдзяйсьняюць хуткія й магутныя рухі, але з хуткім надыходам стомленнасьці, падтрымліваючы толькі кароткія, анаэробныя воплескі актыўнасьці, а наступныя скарачэньні цягліцаў становяцца хваравітымі, выклікаючы пачуцьцё болю. Яны робяць найбольшы ўнёсак у цягліцавую сілу й маюць большы патэнцыял для павелічэньня масы.
Гладкія цягліцы
[рэдагаваць | рэдагаваць крыніцу]Гладкія цягліцы ці «міжвольныя цягліцы» знаходзіцца ў сьценках органаў і структураў, як то страваводу, страўніка, кішачніка, бронхаў, маціцы, мачавыпускальнага канала, мачавой бурбалкі, крывяносных судзінаў. У адрозьненьне ад шкілетных цягліцаў, гладкія цягліцы не знаходзяцца пад сьвядомым кантролем. Гладкія цягліцы складуюцца з характэрных цягліцавых вузаў (міяцытаў). Кароткія пачынакпадобныя вузы гладкіх цягліцаў утвараюць плястыны. Скарачаюцца яны павольна й рытмічна пры падпарадкаваньні сыгналам вэгетатыўнай нэрвовай сыстэмы. Павольныя й працяглыя іхныя скарачэньні адбываюцца міжволі, незалежна ад жаданьня чалавека.
Міякард
[рэдагаваць | рэдагаваць крыніцу]Сардэчная цягліца ці, таксама, зьяўляецца «міжвольнай цягліцай», але яна больш падобная па структуры да шкілетных цягліцаў (амаль ідэнтычная), і знаходзіцца толькі ў сэрцы, і складаецца з кардыяміяцытаў. Скарачэньні сардэчнай цягліцы не падкантрольныя сьвядомасьці чалавека, яна інэрвуецца вэгетатыўнай нэрвовай сыстэмай, гэтак жа як і гладкія цягліцы. Важкую ролю ў інэрвацыі кардыяміяцытаў гуляе кіроўца рытму.
Гістагенэз
[рэдагаваць | рэдагаваць крыніцу]
Усе цягліцы разьвіваюцца з параксіяльнай мэзадэрмы[8]. Параксіяльная мэзадэрма падзяляецца па ўсёй даўжыні зародка на соміты, адпаведна сэгмэнтуючы цела[8]. Кожны сэгмэнт складаецца з трох частак, то бок склератомы (якія ўтварае хрыбетнік), дэрматомы (якая ўтварае скуру) і міятомы (якая ўтварае цягліцы)[8]. Міятома падзелена на дзьве часткі, гіпамэр і эпімэр, якія ўтвараюць разьмешчаныя сьпераду й ззаду ад вэртыкальнай восі цела цягліцы адпаведна[8]. Эпаксіяльныя цягліцы ў чалавека ўтвараюць толькі цягліцу, што трымае хрыбетнік і малыя міжхрыбетныя цягліцы, а таксама інэрвуюцца на сьпінавыя галіны млечавых нэрваў[8]. Усе іншыя цягліцы, у тым ліку цягліцы канцавінаў, месьцяцца сьпераду ад вэртыкальнай восі цела й утвараюцца з вэнтралятэральнымі частымі міятомамі, інэрвуючыся на брушныя галіны млечавых нэрваў[8].
Гісталёгія
[рэдагаваць | рэдагаваць крыніцу]
Шкілетныя цягліцы абабітыя жорсткім плястам злучальнай тканкі, якая называецца эпімізіюмам[9]. Эпімізіюм цягліцаў мацуецца да сухажыльляў да кожнага бока, дзе эпімізіюм становіцца больш тоўстым і калягенавым. Ён таксама абараняе цягліцы ад трэньня ў адносінах да іншых цягліцаў і костак. Эпімізіюм складаецца зь некалькіх пучкоў, кожны зь якіх зьмяшчае ад 10 да 100 і больш цягліцавых валокнаў. Такія пучкі атачае пэрымізіюм, які зьяўляецца шляхам да нэрваў і забясьпечвае прыток крыві ў цягліцы[9]. Ніткападобныя валокны цягліцаў зьяўляюцца асобным відам вузаў (міяцыты), кожная зь якіх захоўваецца ў сваім эндамізіюме калягенавых валокнаў[9]. Такім чынам, агульная сіла складаецца з валокнаў (вузаў), зьвязаных у пучкі, якія самі групуюцца ў цягліцах. На кожным узроўні камплектацыі, калягенавыя мэмбраны атачаюць расслаеньне, і гэтыя мэмбраны дапамагаюць цягліцам падтрымліваць такія функцыі, як то захаваньне ад пасіўнага расьцяжэньня тканкі й распаўсюд сілаў, прыкладзеных да цягліцы[9].
Такія ж сувязі ўзроўняў маюць месца ў будове цягліцавых вузаў, якія складаюцца зь міяфібрылаў, якія самі па сабе зьяўляюцца пучкамі бялковых нітак. Тэрмін «міяфібрылы» ня варта блытаць зь «цягліцавымі валокнамі», якія зьяўляюцца проста іншай назвай для цягліцавых вузаў. Міяфібрылы, што складаюцца зь нітак некалькіх відаў бялковых злучэньняў, арганізаваныя разам у паўтаральныя элемэнты, званыя саркамэрамі. Папярочна-паласатая структура шкілетных і сардэчнай цягліцаў зьяўляецца вынікам заканамернасьці шаблёна разьмяшчэньня саркамэраў у сваіх камэрах. Незважаючы та тое, што абодва гэтыя тыпы цягліцаў зьмяшчаюць саркамэры, валокны ў сардэчнай цягліцы, як правіла, утвараюць больш разгалінаваную сетку. Сардэчныя цягліцавыя валокны злучаныя між сабой інтэркаляванымі дыскамі[10], што надае тканцы зьнешні выгляд сынцыцыюса.
Ніткі саркамэраў складаюцца з скарачальных бялкоў актына й міязіна.
Макраскапічная анатомія
[рэдагаваць | рэдагаваць крыніцу]Макраскапічная анатомія цягліцаў зьяўляецца найбольш важным паказьнікам іхнай ролі ў арганізьме. У большасьці цягліцаў усе валокны арыентаваныя ў тым жа кірунку, у якім звычайна выконваецца іхны рух. У пэнатных цягліцах, асобныя валокны арыентаваныя пад кутом у адносінах да лініі дзеяньня. Праз тое, што валокны такіх цягліцаў цягнуцца пад кутом да агульнага свайго дзеяньня, зьмены цягліцы ў даўжыні меншыя, але гэтая ж арыентацыя дазваляе мець больш валокнаў (і, такім чынам, больш сілы) у цягліцах зададзенага памеру. Пэнатныя цягліцы маюцца, як правіла, у той частцы арганізма, дзе іхная зьмена ў даўжыні менш важная, чым максымальныя высілкі, як то прамая цягліца сьцягна.
Жорсткі, кудзелісты эпімізіюм шкілетных цягліцаў падлучаны да сухажыльляў. У сваю чаргу, сухажыльлі падлучаныя да плясту накастоўя, якое атачае костку й дазваляе ажыцьцяўляць перадачу намаганьняў ад цягліцаў да шкілету. Гэтыя кудзелістыя плясты, разам з сухажыльлямі й зьвязкамі, утвараюць глыбокія фасцыі цела.
Фізыялёгія
[рэдагаваць | рэдагаваць крыніцу]Існуе тры тыпы цягліцаў (шкілетныя, сардэчная й гладкія), якія маюць істотныя адрозьненьні ў будове. Тым ня менш, усе тры тыпы выкарыстоўваць рух актына супраць міязіна для стварэньня сьціску. У шкілетных цягліцаў скарачэньне стымулюецца электрычнымі імпульсамі, якія перадаюцца праз нэрвы, у прыватнасьці праз рухальныя нэрвы. Скарачэньні сардэчнай і гладкай цягліцаў стымулююцца ўнутранымі вузамі, што задаюць рэгулярны тэмп, і распаўсюджваюцца скарачэньнямі ў іншых вузах, якія зьяўляюцца суседнімі ў адносінах да іх. Усе шкілетныя цягліцы й шмат зь якіх гладкіх цягліцаў маюць скарачэньні, якія зьяўляюцца вытворнымі ад нэўрамэдыятара ацэтылхаліну.
Нэрвовы кантроль
[рэдагаваць | рэдагаваць крыніцу]
Эфэрэнтная частка
[рэдагаваць | рэдагаваць крыніцу]Эфэрэнтная частка пэрыфэрыйнай нэрвовай сыстэмы адказвае за транспартаваньне каманды да цягліцаў і залозаў, і нясе поўную адказнасьць за добраахвотны рух. Нэрвы прымушаюць да руху цягліцы ў адказ на добраахвотныя й вэгетатыўныя (міжвольныя) сыгналы ад галаўнога мозгу. Глыбокія цягліцы, павярхоўныя цягліцы, цягліцы твару й унутраныя цягліцы атрымоўваюць сыгналы вылучанай вобласьці ў першаснай маторнай кары галаўнога мозгу, якая разьмяшчаецца непасрэдна наперадзе цэнтральнай разоры, якая дзеліць лобную й цемянную долі.
Акрамя таго, цягліцы рэагуюць на раздражняльнікі рэфлексіўных нэрваў, якія не заўсёды пасылаюць сыгналы да мозгу. У гэтым выпадку сыгнал ад афэрэнтнага валакна не даходзіць да мозгу, але стварае рэфлексіўны рух па прамой сувязі з эфэрэнтнымі нэрвамі ў хрыбетніку. Тым ня менш, большасьць зь цягліцавай дзейнасьці зьяўляецца валявой, то бок ажыцьцяўляецца ў выніку складанага ўзаемадзеяньня паміж рознымі абласьцямі мозгу.
Нэрвы, якія кіруюць шкілетнай мускулатурай у сысуноў, адпавядаюць групам нэўронаў у першаснай маторнай кары галаўнога мозгу. Каманды накіроўваюцца праз базальныя гангліі й мадыфікуюцца на ўваходзе ў мозачак й рэтрансьлююцца празь пірамідны тракт ў млеч, а адтуль да нэрвова-цяглічнага сынапсу й сканчваюцца ў цягліцах. Зваротная сувязь сыгналаў спрыяе падтрыманьню тонусу цягліцаў.
Больш глыбокія цягліцы кіруюцца ядрамі ў ствале мозгу й базальнымі гангліямі.
Афэрэнтная частка
[рэдагаваць | рэдагаваць крыніцу]Афэрэнтная частка пэрыфэрычнай нэрвовай сыстэмы адказвае за транспартаваньне сэнсарнай інфармацыі ў галаўны мозаг, перш за ўсё ад органаў пачуцьцяў, як то скура. У цягліцах, цяглічны пачынак перадае інфармацыю аб ступені даўжыні цягліцы й расьцягнутасьці да цэнтральнай нэрвовай сыстэмы для аказаньня дапамогі ў падтрыманьні паставы й агульнай пазыцыі. Пачуцьцё цела ў прасторы называецца прапрыяцэпцыяй, што зьяўляецца несьвядомым успрыманьнем цела, то бок дасьведчанасьцю аб пазыцыі розных частак цела ў любы момант часу. Гэта можа быць прадэманстравана з дапамогай заплюшчаных вачэй і маханьня рукамі. Мяркуецца, што з дапамогай прапрыяцэптыўнай функцыі, сьвядомасьць чалавека ня будзе губляць веданьне, дзе рукі знаходзяцца на самой справе, нават калі іхнае месцазнаходжаньне не выяўляецца ні адным зь іншых органаў пачуцьцяў.
Некаторыя вобласьці ў мозгу каардынуюць рух і становішча цела з выкарыстаньнем зваротнай сувязі атрыманай ад прапрыяцэпцыі. Мозачак і чырвонае ядро, у прыватнасьці, бесьперапынна вызначаюць пазыцыю ў руху і ўносяць дробныя выпраўленьні дзеля забесьпячэньня ягонай плыўнасьці.
Эфэктыўнасьць
[рэдагаваць | рэдагаваць крыніцу]Эфэктыўнасьць чалавечых цягліцаў, у кантэксьце веславаньня на байдарках і язды на ровары, вагаецца ад 18% да 26%. Гэты паказьнік вызначаецца як стаўленьне выніковай мэханічнай працы да агульнага мэтабалічнага кошту, які можна вылічыць паводле спажываньня кіслароду. Такая нізкая эфэктыўнасьць зьяўляецца вынікам каля 40% эфэктыўнасьці генэрацыі АТФ зь ежы ў энэргію, а таксама страты пры пераўтварэньні энэргіі АТФ у мэханічную працу ўнутры цягліцаў і мэханічныя страты ўнутры цела. Два апошнія віды стратаў залежаць ад тыпу практыкаваньняў і тыпу цягліцавых валокнаў, якія выкарыстоўваюцца пры руху (хуткаскарачальныя або павольныя). Для агульнай эфэктыўнасьці ў 20 адсоткаў, адзін ват мэханічнай энэргіі эквівалентны страце 4,3 ккал у гадзіну. Мэханічная энэргія цыклічнага скарачэньня можа залежаць ад шматлікіх фактараў, у тым ліку актывацыі часу, траекторыі цягліцавай напругі, а, таксама, тэмпаў росту й спадаў сілаў.
Здароўе
[рэдагаваць | рэдагаваць крыніцу]Людзі генэтычна схільныя да перавагі аднаго тыпу цягліцавых групаў над іншымі. Чалавек, які нарадзіўся зь вялікім адсоткам цягліцавых валокнаў першага тыпу тэарэтычна можа больш падыходзіць да такіх відаў спорту, як то трыятлён, хада на адлегласьць і працяглыя гонкі на ровары, у той час як чалавек, які нараджаецца зь вялікім адсоткам валокнаў другога тыпу цягліцаў мае большы шанец атрымаць посьпех у анаэробнай дзейнасьці, як то бег на 200 мэтраў альбо выступы ў цяжкай атлетыцы.
Практыкаваньні
[рэдагаваць | рэдагаваць крыніцу]
Фізычныя практыкаваньні часьцяком раяць як сродак паляпшэньня рухальных навыкаў, падтрыманьня дзеяздольнасьці цягліцаў, трываласьці костак і функцыі суставаў. Практыкаваньні маюць узьдзеяньне на цягліцы, злучальную тканку, костку й нэрвы, якія стымулююць працу цягліцаў. Адным з такіх эфэктаў зьяўляецца цягліцавая гіпэртрафія, то бок павелічэньне ў памерах, што выкарыстоўваецца ў культурызме.
Розныя практыкаваньні патрабуюць перавагі пэўнага выкарыстаньня цягліцавых валокнаў над іншымі. Аэробныя практыкаваньні ўлучаюць у сябе доўгія паводле часу ўзьдзеяньні, зь нізкім узроўнем напружаньня, пры якім цягліцы выкарыстоўваюцца пры значна ніжэйшай іхнай максымальнай сіле скарачэньняў на працягу доўгага пэрыяду часу, самы клясычны прыклад такога практыкаваньня ёсьць маратон. Аэробныя практыкаваньні, якія ў асноўным належаць да аэробнай (з выкарыстаньнем кіслароду) сыстэмы, выкарыстоўваюць больш высокі адсотак цягліцавых валокнаў першага тыпу й спажываюць сумесь тлушчаў, бялкоў і вугляводаў для атрыманьня энэргіі, а таксама вялікую колькасьць кіслароду й амаль не вядуць да вылучэньня малочнай кісьлі.
Анаэробныя практыкаваньні ўлучаюць у сябе кароткія ўспышкі зь вялікай інтэнсіўнасьцю скарачэньняў на значна большы адсотак ад максымальнай сілы сьціску. Прыкладамі анаэробных практыкаваньняў ёсьць бег і ўзьняцьцё цяжараў. Анаэробныя сыстэмы дастаўкі энэргіі выкарыстоўваюцца пераважна цягліцавымі валокнамі другога тыпу альбо хуткаскарачальнымі цягліцамі, яны абапіраюцца ў асноўным на АТФ альбо глюкозу ў якасьці крыніцы энэргіі й спажываюць адносна мала кіслароду, бялкоў і тлушчаў, аднак прыводзяць да ўтварэньня вялікай колькасьці малочнай кісьлі. Практыкаваньні такога тыпу адрозьніваюцца меншым часам актыўнасьці ў параўнаньні з аэробнымі практыкаваньнямі. Многія практыкаваньні й віды спорту маюць спалучэньні аэробных і анаэробных тыпаў актыўнасьці, як то футбол.
Прысутнасьць малочнай кісьлі выклікае тармажэньне ўтварэньня АТФ у цягліцах, але не прыводзіць да стомленасьці, аднак яна можа нават спыніць выпрацоўкі АТФ, калі ўнутрывузавая канцэнтрацыя кісьлі становіцца занадта высокай. Тым ня менш, доўгатэрміновыя практыкаваньні выклікаюць нэаваскулярызацыю ў цягліцах, што павялічвае здольнасьць вываду рэчываў зь цягліцаў і падтрымлівае іхны сьціск. Малочная кісьля можа быць выкарыстаная іншымі цягліцамі або тканкамі арганізма ў якасьці крыніцы энэргіі, або яна транспартуецца ў печань, дзе ператвараецца назад у піруват. У дадатак да павышэньня ўзроўню малочнай кісьлі, фізычныя нагрузкі прыводзяць да страты іёнаў каля ў цягліцах і прыводзяць да павелічэньня канцэнтрацыі іёнаў каля недалёка ад цягліцавых валокнаў, у інтэрстыцыіюме. Падкісьленая малочная кісьля можа прывесьці да аднаўленьня сілаў, такім чынам, што ацыдоз можа абараніць ад стомы, а не быць прычынай стомленасьці[11].
Гіпэртрафія
[рэдагаваць | рэдагаваць крыніцу]Незалежна ад сілы й паказьнікаў, цягліцы могуць расьці большымі праз цэлы шэраг фактараў, у тым ліку парушэньне выпрацоўкі гармонаў, фактары разьвіцьця, наяўнасьць сілавых трэніровак і хваробы. Насуперак распаўсюджанаму меркаваньню, колькасьць цягліцавых валокнаў ня можа быць павялічаная з дапамогай практыкаваньняў. Замест гэтага, цягліцы растуць за кошт спалучэньня росту цягліцавых вузаў у новыя бялковыя злучэньні, якія спрыяюць дадатковай масе й зьяўляюцца недыфэрэнцыяванымі вузамі, якія знаходзяцца разам з існымі цягліцавымі вузамі[12]. Цягліцавыя валокны маюць абмежаваныя магчымасьці для росту за кошт гіпэртрафіі.
Біялягічныя фактары, як то ўзрост і ўзровень гармонаў могуць уплываць на цягліцавую гіпэртрафію. У пэрыяд плоцевага пасьпяваньня ў мужчынаў, гіпэртрафія адбываецца ў паскораным тэмпе, бо тэмпы росту стымулявальных гармонаў, што выпрацоўваюцца арганізмам, павялічваюцца. Прыродная гіпэртрафія звычайна спыняецца ў канцы падлеткавага ўзросту. З прычыны таго, што тэстастэрон зьяўляецца адным з асноўных гармонаў росту арганізма, у сярэднім, мужчыны дамагаюцца большай гіпэртрафіі, чым жанчыны. Прымаючы дадатковы тэстастэрон і іншыя анабалічныя стэроіды, можна павялічыць цягліцавую гіпэртрафію.
Атрафія
[рэдагаваць | рэдагаваць крыніцу]
Бязьдзейнасьць і голад у сысуноў можа прывесьці да атрафіі шкілетных цягліцаў, зьніжэньню цягліцавай масы, што можа суправаджацца меншым лікам і памерам цяглічных вузаў, а таксама зьніжэньню ўтрыманьня бялку[13]. Цягліцы атрафуюцца, таксама, як вынік працэсу натуральнага старэньня ці ад хваробы.
У людзей, якія доўгі час правялі ў нерухомасьці, як то ў выпадку ложкавага рэжыму альбо палёту ў космас, як вядома, назіраецца паслабленьне цягліцаў і іхная атрафія. Атрафія ўяўляе асаблівую цікавасьць для пілятуемых касьмічных палётаў, бо досьвед бязважкасьці пры касьмічных палётах зьяўляецца чыньнікам страты да 30% масы некаторых цягліцаў[14][15]. Такія наступствы адзначаюцца таксама пры сьпячцы ў малых сысуноў, як то ў карычневых кажаноў[16].
Падчас старэньня, адбываецца паступовае зьніжэньне здольнасьці падтрымліваць функцыі шкілетных цягліцаў і масы. Гэтая зьява вядомая як саркапэнія. Дакладная прычына саркапэніі невядомая, але яна можа быць зьвязаная са спалучэньнем паступовага замаруджваньня выпрацоўкі «спадарожнікавых вузаў», якія дапамагаюць рэгенэрацыі валокнаў шкілетных цягліцаў, зьніжэньня адчувальнасьці. Саркапэнія зьяўляецца нармалёвым аспэктам старэньня, і на самой справе ня лічыцца хваравітым станам, але яна можа быць зьвязана са шматлікай колькасьцю траўмаў у пажылых людзей, а таксама зьніжэньнем якасьці жыцьця[17].
Ёсьць таксама шмат захворваньняў і станаў, якія выклікаюць атрафію цягліцаў. Іхнымі прыкладамі зьяўляюцца рак і СНІД, якія выклікаюць сындром зьнясіленьня цела, вядомы як кахексія. Іншымі сындромамі або ўмовамі, якія могуць выклікаць атрафію шкілетных цягліцаў, ёсьць застойная хвароба сэрца й некаторыя захворваньні печані.
Захворваньні
[рэдагаваць | рэдагаваць крыніцу]
Нэрвова-цягліцавыя захворваньні ўплываюць на цягліцы або іхны нэрвовы кантроль. Увогуле, праблемы з нэрвовай рэгуляцыяй могуць прывесьці да паралічу або спастычнасьці, у залежнасьці ад месца й характару праблемы. Большая частка нэўралягічных разладаў, пачынаючы ад парушэньня мазгавога кровазвароту (інсульт) і хваробы Паркінсона й сканчаючы хваробай Кройтцфэльта—Якаба, можа прывесьці да праблемаў з рухам або каардынацыяй рухаў цягліцаў.
Сымптомы захворваньня цягліцаў могуць улучаць слабасьць, спастычнасьць, міяклянію й міяльгію. Дыягнастычныя працэдуры, якія могуць выявіць цягліцавыя разлады ўлучаюць тэставаньне на крэатынкіназу ў крыві й вымярэньне электрычнай актыўнасьці ў цягліцах. У некаторых выпадках можа быць праведзеная біяпсія цягліцаў дзеля выяўленьня міяпатыі, а таксама генэтычнае тэставаньне для выяўленьня адхіленьняў у ДНК, зьвязаныя са спэцыфічнымі відамі міяпатыі й дыстрафіі.
Неінвазіўная элястаграфічная тэхніка, якая вымярае шумы ў цягліцах, зараз праходзіць выпрабаваньне дзеля забесьпячэньня магчымасьці маніторынгу нэрвова-цягліцавых захворваньняў. Гук, выраблены цягліцамі, вылучаецца ад скарачэньняў нітак актаміязіна ўздоўж восі цягліцаў. Падчас скарачэньня, цягліца скарачаецца ўздоўж сваёй падоўжнай восі й пашыраецца ў папярочнай восі, вырабляючы вібрацыі на паверхні[18].
Цягліцы чалавека
[рэдагаваць | рэдагаваць крыніцу]-
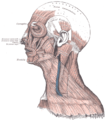 Цягліцы шыі, від збоку
Цягліцы шыі, від збоку -
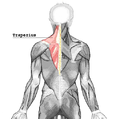
 Цягліцы тулава, від ззаду
Цягліцы тулава, від ззаду -
-

-
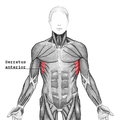
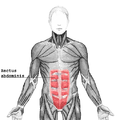
 Цягліцы тулава, від сьпераду
Цягліцы тулава, від сьпераду -

-
-
-

-

-
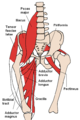
-
 Цягліцы нагі (від ззаду)
Цягліцы нагі (від ззаду) -
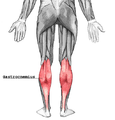
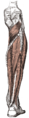 Цягліцы галёнкі, від ззаду
Цягліцы галёнкі, від ззаду -

-














Крыніцы
[рэдагаваць | рэдагаваць крыніцу]- ^ Цягліца. Slounik.org. Праверана 16 лютага 2013 г.
- ^ Літаратурны тлумачальны слоўнік. Родныя вобразы. Праверана 16 лютага 2013 г.
- ^ Анатоль Леанцюк Мышцы // Беларуская энцыклапедыя ў 18 тамах / Генадзь Пашкоў. — Менск: Беларуская энцыкляпэдыя, 2000. — Т. 11. — С. 54. — 560 с. — 10 000 ас. — ISBN 985-11-0188-5
- ^ Julie McDowell. Encyclopedia of Human Body Systems. 2 volume. — Greenwood, 2010. — С. 326. — ISBN 978-0-3133-9175-0
- ^ Marieb, EN; Hoehn, Katja. Human Anatomy & Physiology. — 8th ed. — San Francisco: Benjamin Cummings, 2010. — 312 с. — ISBN 978-0-8053-9569-3
- ^ а б McCloud, Aaron. (2010) «Build Fast Twitch Muscle Fibers». Complete Strength Training. Праверана 6 лютага 2013 г.
- ^ Larsson, L; Edström, L; Lindegren, B; Gorza, L; Schiaffino, S. «MHC composition and enzyme-histochemical and physiological properties of a novel fast-twitch motor unit type». — The American Journal of Physiology 261 (1 pt 1). — С. 93–101.
- ^ а б в г д е Sweeney, Lauren. Basic Concepts in Embryology: A Student's Survival Guide. — (1st Paperback ed.). — McGraw-Hill Professional, 1997.
- ^ а б в г MacIntosh, BR; Gardiner, PF; McComas, AJ. 1. Muscle Architecture and Muscle Fiber Anatomy // Skeletal Muscle: Form and Function. — (2nd ed.). — Champaign, IL: Human Kinetics, 2006. — С. 3–21. — ISBN 0-7360-4517-1
- ^ Kent, George C. 11. Muscles // Comparative Anatomy of the Vertebrates. — (7th ed.). — Dubuque, Iowa, USA: Wm. C. Brown Publishers, 1987. — С. 326–374. — ISBN 0-697-23486-X
- ^ Nielsen, OB; de Paoli, F; Overgaard, K. «Protective effects of lactic acid on force production in rat skeletal muscle». — Journal of Physiology 536 (1). — 2001. — С. 161–166.
- ^ Poole, RM. The Incredible Machine. — Washington, DC: National Geographic Society, 1986. — С. 307–311. — ISBN 0-87044-621-5
- ^ Fuster, G; Busquets, S; Almendro, V; López-Soriano, FJ; Argilés, JM. «Antiproteolytic effects of plasma from hibernating bears: a new approach for muscle wasting therapy?». — Clin Nutr. — 2007 Т. 26 (5). — С. 658–661.
- ^ Roy, RR; Baldwin, KM; Edgerton, VR. «Response of the neuromuscular unit to spaceflight: What has been learned from the rat model». — Exerc. Sport Sci. Rev.. — 1996 Т. 24. — С. 399–425.
- ^ NASA Muscle Atrophy Research (MARES). NASA (2010). Праверана 6 лютага 2013 г.
- ^ Lohuis, TD; Harlow, HJ; Beck, TD. Hibernating black bears (Ursus americanus) experience skeletal muscle protein balance during winter anorexia. — Comp. Biochem. Physiol. B, Biochem. Mol. Biol.. — 2007 Т. 147 (1). — С. 20–28.
- ^ Roche, Alex F. Sarcopenia: A critical review of its measurements and health-related significance in the middle-aged and elderly. — American Journal of Human Biology. — 1994 Т. 6. — С. 33.
- ^ Dumé, Belle (2007) 'Muscle noise' could reveal diseases' progression Праверана 6 лютага 2013 г.
Літаратура
[рэдагаваць | рэдагаваць крыніцу]- Машчанка М. Біялогія: вучэб. дапам. для 9-га кл. устаноў агульн. сярэдн. адукацыі з бел. мовай навучання. — 3-е. — Мн.: Народная асвета, 2004. — ISBN 978-985-03-1531-1 (бел.)
- Гайворонский И.В., Ничипорук Г.И. Анатомия мышечной системы. — СПб.: ЭЛБИ-СПб, 2005. — С. 84. (рас.)
- Миловзорова М. Анатомия и физиология человека. — Медицина, 1972. (рас.)
- Швырев А.А. Анатомия и физиология человека с основами общей патологии. — Феникс, 2004. — 416 с. (рас.)
Вонкавыя спасылкі
[рэдагаваць | рэдагаваць крыніцу]![]() Цягліцы — сховішча мультымэдыйных матэрыялаў
Цягліцы — сховішча мультымэдыйных матэрыялаў
- Сіла чалавечага сэрца. The Physics Factbook. (анг.)
- Даведнік цягліцаў чалавека (анг.)